


NEURÔNIOS
São células extremamente especializadas que, desta forma, apresentam baixíssimo potencial mitótico. Sua morfologia, rica em prolongamentos, possibilita a conexão com os mais variados tipos celulares, pertencentes ou não ao próprio tecido nervoso. Como modelo genérico para um neurônio devemos imaginar uma célula dividida em três porções: dendritos, corpos celular e axônio.
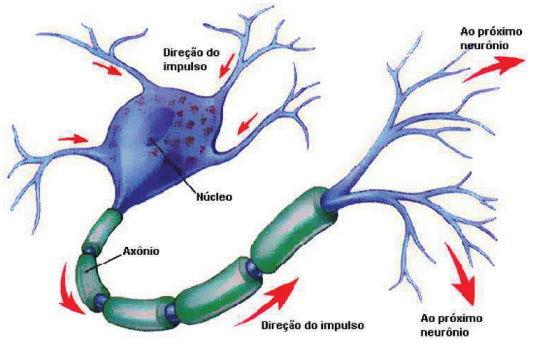
• Dendritos: prolongamentos numerosos, especializados na função de receber os estímulos do meio ambiente, de células epiteliais sensoriais ou de outros neurônios.
• Corpo Celular: região na qual se encontra o núcleo da célula, apresenta elevada capacidade de processamento de informações metabólicas que culminam na síntese de proteínas específicas que ditam o comportamento da célula.

• Axônio: prolongamento único e, em geral, mais comprido, é especializado na condução de impulsos que transmitem informações de um neurônio a outra célula.
• Bainha de mielina: atua como um isolamento elétrico e aumenta a velocidade de propagação do impulso nervoso ao longo do axônio.
TIPOS DE NEURÔNIOS
Nem sempre as regiões de um neurônio são tão bem definidas como no modelo geral descrito anteriormente. Assim, é possível encontrar neurônios com diferentes morfologias que são classificados como: unipolares, bipolares, Pseudounipolares e multipolares

Neurônios unipolares apresentam uma região bem definida de corpo celular, mas não possuem dendritos, sendo seu único prolongamento responsável por uma função axonal. Os neurônios bipolares possuem um corpo celular localizado entre dois prolongamentos, um dendrítico e outro axonal. Neurônios pseudounipolares parecem apresentar um único prolongamento, porém, como este sofre uma bifurcação, uma de suas extremidades atua como dendrito e outra como axônio. Por fim, neurônios multipolares, mais típicos, apresentam um corpo celular rodeado por prolongamentos, sendo a maioria de dimensões reduzidas e função dendrítica, e um único mais comprido de função axonal.
Veremos neste módulo que propagação de um impulso nervoso dependa da troca de íons entre o citoplasma e o meio extracelular. Estes íons acabam por atuar, de uma forma simplificada, como cargas elétricas. Assim, quanto maior for a região do axônio onde estas trocas de cargas elétricas precisem ocorrer, mais tempo o impulso demorará para atingir a extremidade da célula.
É no contexto de classificação dos neurônios quanto à velocidade de propagação do impulso nervoso que reparamos na presença da bainha de mielina. Esta substância de característica lipídica, produzida a partir de células especializadas conhecidas como células de Schwann (sistema nervoso periférico) ou oligodendrócitos (sistema nervoso central), reveste os axônios de maneira descontínua e acaba por “encurtar” a região a ser percorrida pelo impulso. Desta forma, neurônios mielinizados acabam por apresentar maior velocidade na condução de um impulso nervoso quando comparados a neurônios não mielinizados.
Repare que no neurônio mielinizado representado a seguir, as únicas regiões de seu axônio nas quais há trocas de cargas elétricas são os espaços de descontinuidade entre regiões da bainha de mielina, os nódulos de Ranvier. Como o impulso nervoso não percorre todo o axônio de neurônios mielinizados, o caracterizamos como um impulso saltatório, em oposição ao impulso contínuo característico dos neurônios não mielinizados.

CÉLULAS DA GLIA
Como qualquer tipo de tecido, o tecido nervoso também apresenta diferentes tipos celulares. Apesar do grande destaque dado aos neurônios, seu grau de especialização é tão grande que mesmo funções básicas de sobrevivência acabam sendo desempenhadas por células de suporte fisiológico, as células da glia:
• Oligodendrócito: responsável pela síntese da bainha de mielina no sistema nervoso central.
• Célula de Schwann: responsável pela síntese da bainha de mielina no sistema nervoso periférico.
• Microglia: responsável pela defesa do tecido contra patógenos e pela reciclagem de moléculas provenientes de células mortas.
• Astrócito: responsável pela conexão entre neurônios e vasos sanguíneos, possui inúmeros prolongamentos que aumentam sua superfície de contato e viabilizam sua função de nutrição.

TRANSMISSÃO DO IMPULSO NERVOSO
A função de um impulso nervoso é promover a liberação de mensageiros químicos (neurotransmissores) que se encontram em vesículas membranosas no interior do citoplasma dos neurônios. Esta liberação, no entanto, é facilitada pela desorganização da estrutura da membrana através da rápida passagem de íons do meio extracelular para o meio intracelular e vice-versa. É este fluxo de íons que, na verdade, batizamos de impulso nervoso.
Para compreender as bases moleculares de um impulso devemos lembrar que um neurônio, como toda célula em nosso corpo, é polarizado. Isso significa dizer que sua membrana plasmática é carregada positivamente em sua face extracelular e negativamente na face citoplasmática. A diferença de polarização é causada pela variação da concentração de íons de sódio (Na+) e potássio (K+) dentro e fora da célula. Esta diferença estável é mantida a custa do ATP, pois o sódio é forçado a sair da célula enquanto o potássio é forçado a entrar na célula através da bomba de sódio e potássio.

No estado polarizado, o neurônio encontra-se carregado positivamente do lado de fora e negativamente do lado de dentro.
Quando o neurônio é estimulado ocorre a sua despolarização, já que o sódio (Na+) que estava em maior quantidade do lado de fora da célula passa a penetrar o citoplasma através da abertura de canais de sódio. Este influxo de sódio aumenta a quantidade de cargas positivas no interior da célula em relação ao seu lado externo, invertendo os pólos da membrana plasmática. Este fenômeno de despolarização é, em última análise, a própria propagação de um impulso nervoso, uma vez que o movimento rápido e intenso de íons através da membrana a desestabiliza de forma que, ao chegar ao final do axônio, favorece-se a liberação dos neurotransmissores.

Na despolarização, há passagens de íons Na+ para o interior da célula que fica carregada positivamente.
Os canais de sódio, no entanto, permanecem abertos por uma fração mínima de tempo. Conforme canais mais próximos ao término do axônio vão se abrindo, aqueles que ficaram para trás, mais próximos ao corpo celular, vão se fechando. Isto impende que novos íons de sódio entrem na célula e seu retorno ao meio extracelular volta a ocorrer em razão da atividade da bomba de sódio e potássio.
Neste momento, por outro lado, abrem-se os canais de potássio, promovendo o efluxo destes íons ao meio extracelular. Assim, se antes o neurônio polarizado havia sido despolarizado pela entrada de íons de sódio, agora ele é repolarizado com a saída de íons de potássio que objetivam o retorno deste neurônio ao seu estado de repouso.
Similar ao que observamos para os canais de sódio, os canais de potássio também permanecem abertos por pouco tempo. Este tempo é suficiente para que as cargas se re-equilibrem, sendo a concentração de Na+ elevada na face extracelular da membrana plasmática, e a concentração de K+ elevada em sua face intracelular.
Como um impulso nervoso depende da alteração das cargas elétricas de um axônio, é possível medir a diferença de potencial (DDP) existente entre suas faces citoplasmática e extracelular. Esta DDP é da ordem de milivolts (mV) e caracteriza um neurônio em estado de repouso com aproximadamente -60 mV. Durante a abertura dos canais de sódio, a despolarização eleva essa DDP à aproximadamente +40 mV. Por fim, o fechamento dos canais de sódio e abertura dos canais de potássio repolariza o neurônio, como mostrado na figura a seguir.

Dois pontos importantes nesse gráfico devem ser notados. Um deles diz respeito ao limiar de ação, que configura o mínimo de despolarização que um neurônio deve atingir mediante estímulo para que, de fato, um impulso nervoso seja transmitido. Isto garante que estímulos fracos demais não levem a qualquer atividade neuronal. O segundo diz respeito ao momento de repolarização que reduz a DDP a um valor inferior ao potencial de repouso, conhecido como período refratário. A ideia, neste caso, é garantir que não haja o disparo de impulsos nervosos consecutivos, dando tempo para que o organismo reaja. Assim, passado este período refratário, caso seja necessário e a DDP já tenha atingido o potencial de repouso novamente, um novo estímulo pode desencadear uma nova despolarização e propagação de impulso nervoso.
É importante notar que o impulso nervoso propaga-se em apenas uma direção. Ele caminha dos dendritos para o corpo celular e deste para o axônio. Ao chegar na extremidade do axônio, ocorre a liberação dos neurotransmissores em um fenômeno de comunicação conhecido como sinapse.
SINAPSE
A sinapse é fenômeno de comunicação entre um neurônio e outra célula, que pode ser um neurônio ou glândula, por exemplo. Esta comunicação não envolve o contato direto entre as membranas das células envolvidas, mas a liberação de mensageiros químicos em um pequeno espaço conhecido como fenda sináptica de mais ou menos vinte nanômetros. A membrana pela qual são liberados os transmissores se chama membrana pré-sináptica, e a membrana que recebe os neurotransmissores se chama membrana pós-sináptica.

Os neurotransmissores, como mensageiros, ligam-se aos seus receptores específicos na membrana pós-sináptica e estimular a modificação do metabolismo desta célula. Isto pode significar a contração muscular, a aceleração da frequência cardíaca, a secreção de um hormônio, etc. Os neurotransmissores mais conhecidos são a acetilcolina e a noradrenalina mas existem outros como a endorfina e a dopamina, por exemplo.

